A reader asked my opinion about my reported R1b subclade of one low quality sample from Ra’s Al-Ḥamrāʾ 5 necropolis, Muscat (Oman), published (without Y-DNA) in Harney et al. (2020). For those interested, here are the relevant calls, with information on the graves taken from Salvatori (2007):
I11919_I11920_I11921: Grave 221 (ca. 3700-3200 BC), mtDNA H2a2a1, Y-DNA R1b>L754 (xPH155; xL389>P297>M269; xPF6323>PF6292).
* The samples show a straightforward path (but full of deamination question marks): CT (with 1 ancestral call M5813 1x C->A) -> P>P295>M45>P284>P226 -> K>M526>YSC0000186 -> R -> R1b>L754. Ancestral for, among others: A00; A1a; C; D; E; G; H; I; J; LT; L; N; R1a-M459; R1a-YP4141; R2; T.
LOCUS : HSQB/C-HSVA/D
Orientation : NE-SW | F.or. : NW | Position : 4.Y.Lm.Lm. (Crouching, on the right side)
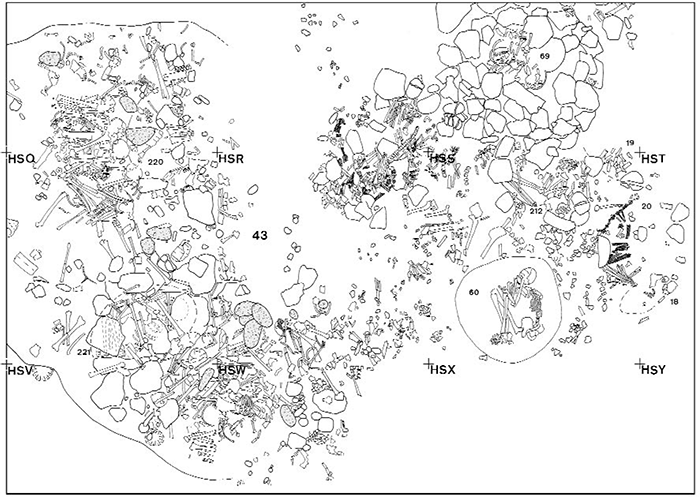
COMMENTS: Massive covering of limestone blocks on which were laid numerous human remains, apparently pertaining to the south-western extension of Area 43. The filling, at various heights, had some infant skeletal remains (part of the cranium and some long bones) probably from a grave disturbed by the excavation of the Grave 221 pit. In the filling were found modest quantities of faunal remains (100g of fish bones and 100g of fragmented land mammal bones). In the uppermost layer of the pit filling were found, evidently sliding in from the surrounding area, a bone pin (Inv.DA 7218: L. 8.2, w 1.1, th 0.3) from capriovine tibia, two Engina mendicaria beads and three cylindrical soapstone beads (Inv.DA 7219: 1.∅ 0.75, H. 0.6; 2. ∅ 0.80, H. 0.59; 3. ∅ 0.75, H. 0.42). On the human skull, under the limestone covering, four peridotite pebbles were lying. The deceased lay directly on the limestone bedrock platform.
ANTHROPOLOGICAL NOTES: State of preservation: fragmentary. Skull and postcranial skeleton are largely incomplete. Average stature is 1685.1 mm.
Area 43. Map of the burial complex at the end of the 1984 campaign (area 43) and grave 221 at the bottom left. Featured image of this post, on the top: Grave 221. Left: PLATE 53.G. 221 stone covering. Right: PLATE 54.G. 221 skeletal remains. Images modified from Salvatori (2007).
* Its simpler path is CT -> F -> P-P226. Ancestral for, among others: A1a, C, D, G, H, IJ, LT, N. Derived for one A1b equivalent (P108, 2x C->T) but nothing below it. The original question was regarding an L23 (G->A) call for this sample, which does not pass a minimum quality threshold, and is contradicted by the better quality call for ancestral M269 of the other sample. Apart from that, R1b-L23 (defined by three SNPs, and with a TMRCA, formation date, and star-like phylogeny starting in the mid- to late-5th millennium) appears to be incompatible with the age and archaeological context of both samples.
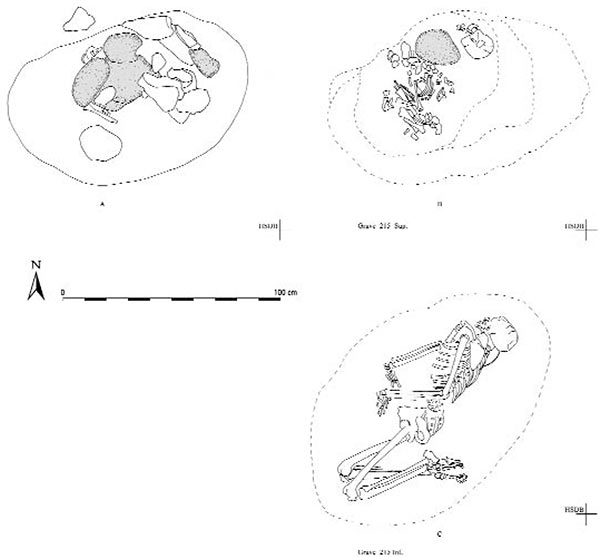
My guess is that this sample refers to the inferior layer of grave 215, which is better preserved than the infant from the superior layer:
LOCUS : HSDB/C
Orientation : NE-SW | Position : 4.Y.Hk.Hk. (Crouching, on the right side).
COMMENTS: Lacking a covering, the grave was identified only by chromatic contrast of the soil at an average depth of 30 cm above the human remains. The pit filling yielded only a considerable quantity of shells (1040 g).The grave was probably disturbed to some measure by the excavation of the Grave 215 sup. pit. Such disturbance also produced a displacement of the deceased’s cranium.
ANTHROPOLOGICAL NOTES: State of preservation: good. Sex: M (M = 1.23, calculated on 20 traits). Age: 39-45 years. Average stature is 1670.2 mm.
FIGURE 165. Plan of G. 215 Sup. (A-B) and G. 215 Inf. (C).
When I first saw these low quality samples, and after looking for the site and reading about “fisher gatherers”, I only considered reliable the better quality one, from the disturbed area 43, and – even assuming that it was R1b, and that it belongs to its supposed archaeological layer – I thought it would be mostly relevant to support a potential southern route of expansion of R1b-L754 from Central Asia through Iran into Europe.
In any case, the ancestral call for xPF6323>PF6292 (1x C->T, also subject to deamination), was enough for me to reject a relationship to the expansion of “African R1b-V88”. Even worse, since these samples were close to Iran and dated to the mid-4th millennium BC – a time when many relevant migrations had already happened elsewhere in the world – it offered hardly any evidence beyond some lost R1b-L754* branch potentially related to the Villabruna cluster, which had spread about 10,000 years earlier.
SNP Tracker‘s estimated migration route for R1b-L754.
On the other hand, now that I have rechecked the second individual, from grave 215, it looks like both males – through SNPs obtained from three samples each – point (through ancestral and derived calls) to an R1b subclade, and that they belong (based on different burial grounds) to the same archaeological and chronological layer.
Relevant excerpts (emphasis mine, stylistic changes for clarity):
It is important to stress here that what is called “Neolithic” in Eastern Arabia is clearly different from the Neolithic in the Near East and has followed its own path (Cleuziou, 2005). In Eastern Arabia, this period is characterized by an absence of agriculture. Oasis agriculture, based on palm tree cultivation (Phoenix dactylifera), is not attested until 3000 BC in the region (Tengberg, 2012), and characterizes the beginning of the Bronze Age. Herding (sheep, goat, cattle) was practiced since 5500 BC in the UAE and Oman in combination with hunting, gathering, and an intensive exploitation of marine resources on the coast (Uerpmann, 2003). Neolithic populations are thought to have had a nomadic or semi-nomadic way of life, although the patterns of mobility remain to be investigated and some degree of sedentarity may have been experienced at a few sites (Biagi and Nisbet, 2006).
The issue of seasonality of site occupation has been debated for RH-5 as for the other Neolithic sites in the area. Several authors have interpreted the occupation of RH-5 as seasonal. Seasonal mobility of the human groups has been proposed based on several indirect arguments, including the fragmentation and the seasonal availability of natural resources (Tosi, 1986; Uerpmann, 2003), the preferential orientation of the deceased, and the evaluation of the size of the population (Coppa et al., 1985; Salvatori, 2007; Cavulli and Scaruffi, 2013). Other authors have considered the possibility of a more permanent occupation of the site. Macchiarelli (1989) observed a reduced dental size in the RH-5 population, interpreted as a mark of genetic isolation and increasing population density, suggesting a sedentary or semi-sedentary lifestyle. The presence of young domestic animals led Uerpmann (2003) to hypothesize that at least part of the group was living at RH-5 during most of the year. Fruits of Ziziphus and seeds of Setaria were also recovered at RH-5, suggesting the presence of part of the group at least during the summer.
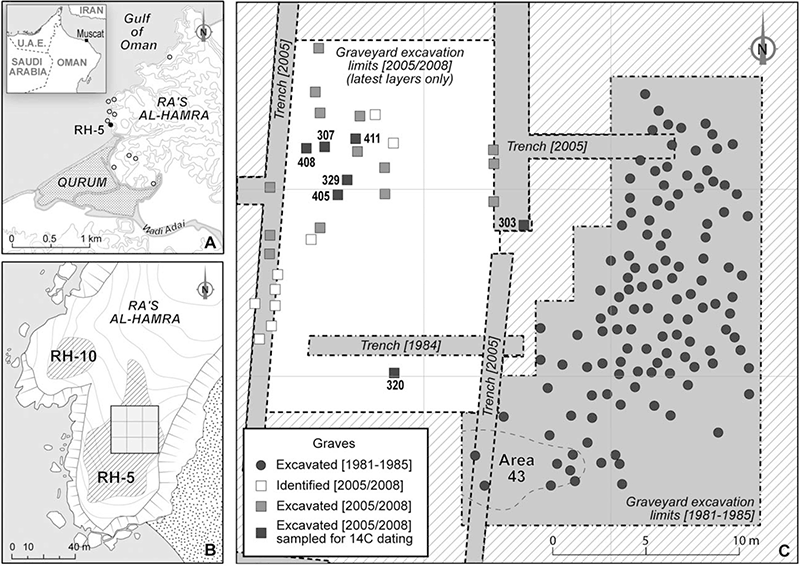
(A) Location of RH-5 and other prehistorical sites in the area of Ra’s al-Hamra and Qurum (Muscat, Oman). (B) Closeup on the Ra’s al-Hamra promontory; the square indicates the location of the RH-5 graveyard. (C) Location of the graves identified (white squares), excavated (gray dots and squares) and sampled for the study of Zazzo, Muñoz, & Saliège 2013 (black squares).
The analysis of the settlement structures, material culture assemblages, and handicraft activities of several 5th millennium BP shell middens of the coast of Oman led Biagi and Nisbet (2006) to the conclusion that “a more sedentary way of life” had already taken place in this area among the Middle Holocene fisher-gatherers. The authors suggest a “relative permanence of their inhabitants throughout a long period of the year, at least during the autumn and winter months.” These authors did not go as far as postulating a permanent, all year round, occupation for the site. The low stable isotope variability (less than 2‰), coupled with relatively high δ13C values, suggests that immigrants from the hinterland are absent from our sample, in keeping with the high degree of endogamy postulated for the group (Coppa et al., 1985, 1990; Macchiarelli, 1989; Coppa and Cucina, 2007). Given the dominance of the marine signal in the diet, it is unlikely that the individuals analyzed here were leaving the coast for long periods of time (i.e., 6 months). We cannot however, exclude mobility of part or all of the population along the coast. (…)
The climatic conditions of the Arabian Peninsula started to deteriorate around the end of the 6th millennium BC (Sanlaville, 1992) and this lead to a drastic decrease in the number of sites in the interior and on the coast of the Persian Gulf at the end of the 5th millennium BC (Uerpmann, 2003). In contrast, the sites located on the Omani coast seemed to flourish during the 4th millennium BC, when today’s arid conditions had almost been established (Berger et al., 2013). Most of these sites installed on the Omani coast are located close to the mouth of wadis where permanent sources of freshwater and highly productive marine environments were accessible. Several of them (Ra’s al-Hamra RH-5/6/ 10, Wadi Shab GAS-1) have yielded graveyards used over generations, reflecting the anchoring of populations in these places. These sites could be interpreted as refuges where Mid-Holocene populations could have experienced sedentarization.
At the end of the 4th millennium BC, the societies occupying the peninsula of Oman underwent profound economic changes with the entry into a network of inter-regional trade, the adoption of new technologies (metallurgy, pottery), new modes of food production (oasis agriculture, irrigation, intensification of fishing), and innovations in conservation techniques of agriculture products and fisheries (Cleuziou, 2002). These innovations were accompanied by social changes, particularly noticeable through changes in settlement patterns (permanent structures of mud brick), and burial practices (Cleuziou and Munoz, 2007; Méry and Charpentier, 2009). This is reflected by an increase in population and a change of occupation and land use.
Oman BA samples
Because these samples offer the only ancient Y-DNA haplogroups we have from Arabia, and are important to understand the expansion of the Neolithic – and thus potentially of Afroasiatic languages (cf. Prendergast et al. 2019) – my guess is that their Y-SNPs will be much more discussed than many other similar low-quality European ones with questionable (terminal) calls; or than they would if a similar call were made of, say, hg. E or J.
However, if we disregard for a moment the questionable reliability of their derived SNPs, assuming that most samples from the region are going to have a similarly low quality, and accepting that all combined SNPs from both individuals support each other in pointing to R1b(xM269), the speculative discussion below about the spread of R1b-PF6323 becomes much more interesting.
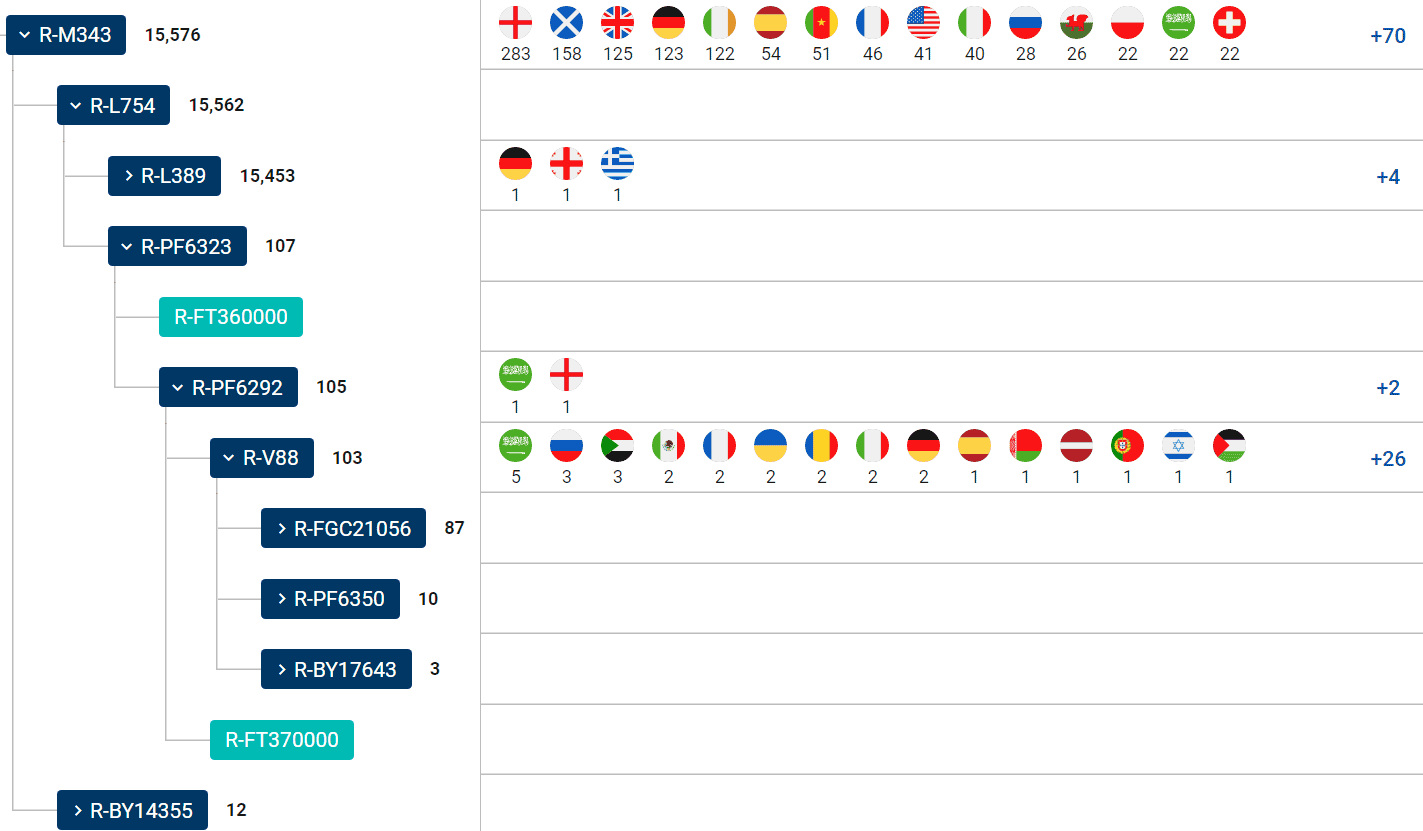
This warrants first a closer look at the current state of the FamilyTreeDNA Haplotree, with a revamped PF6323 branch including ancient samples, thanks to the efforts of the team formed by phylogeneticist Michael Sager and Göran Runfeldt from the R&D team. Together with their modern distribution, the attestation of each branch in ancient samples offers a clue slightly different to what we had until now, one more relevant for potential Neolithic migration waves.
NOTE. YFull’s V2219 tree is not only clearly lagging behind – defining only two levels, V2219 and V88, after a year of being cognizant of the new tree. There are also some inconsistently defined equivalents, like PF6314 for V2219, when it is part of V88 in FTDNA. In fact, this SNP is supposed to be present in both V2219* samples in their tree, OC1_Meso and SC2_Meso, which are negative for V88.
Capture of FTDNA Haplotree’s R-PF6323 branch as of 30th January 2021. Notice modern Saudi testers on the upper clades, as well as interspersed African and European ones. Isolated branches defined by ancient samples in light blue.
The reason why I discarded at first the relevance of these Oman BA samples for the history of R1b-PF6323 is that I had previously (2018) associated the migration of the R1b-V88 branch with a Mesolithic route through Italy into Northern Africa and subsequent bottlenecks through the Green Sahara. I changed later (2020) to a Neolithic spread through the Central Mediterranean, when a more accurate split of ancient clades was proposed, coupled with the finding of Neolithic ancestry spreading with East African pastoralists. Because of this, it did not make sense to find some potentially basal R1b-PF6323* in a population that should have been strongly bottlenecked that far to the east, well below R1b-V88.
However, this reasoning was based on the assumption that the sample was xPF6323>PF6292, which is not clear; on the nature of this population as indigenous fisher-gatherers, which was too simplistic; and on two concepts about the traditional “V88” tree and distribution that in hindsight seem weak:
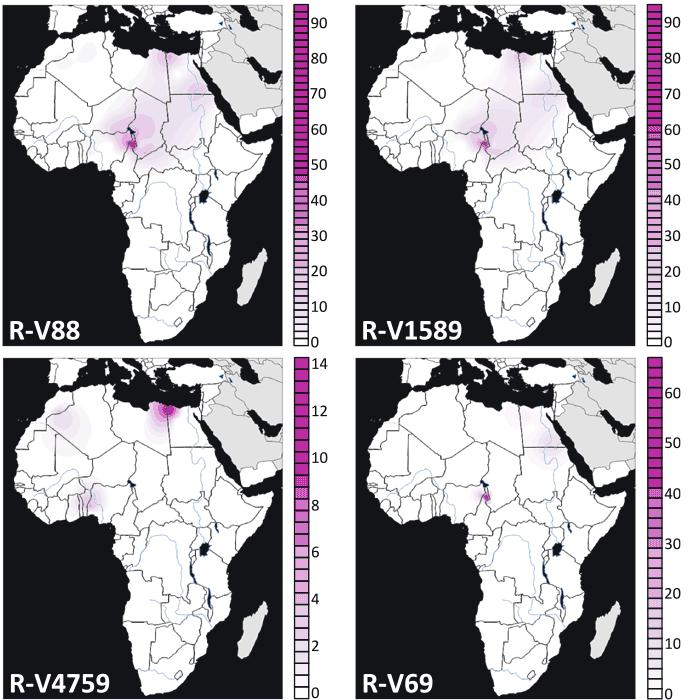
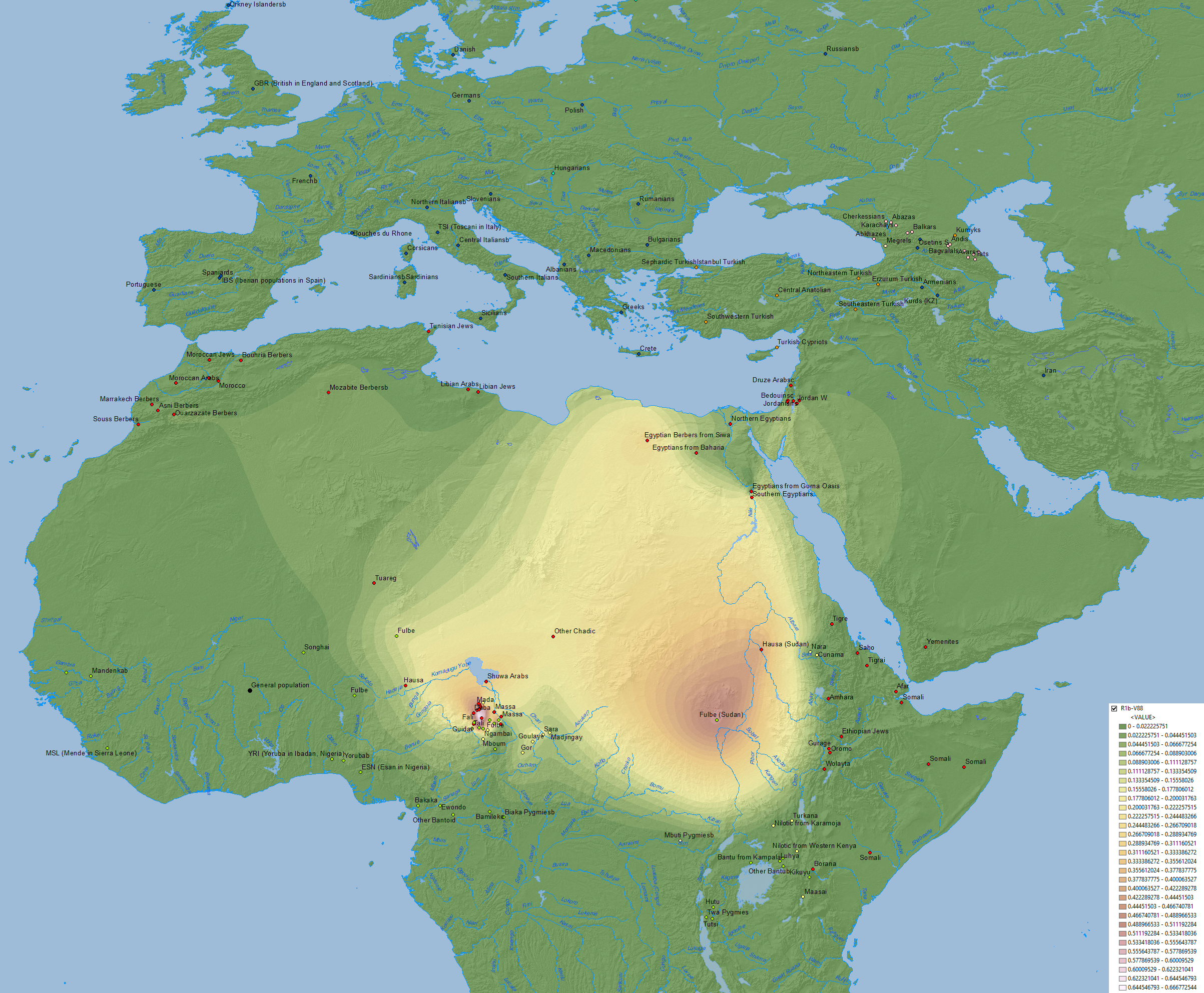
1) The phylogeographic assessment in D’Atanasio et al. (2018), based on the older R1b-V88 tree, which potentially includes branches above it (PF6323 and PF6323>PF6292) and other branches stemming from those (PF6323>FT360000 and PF6323>PF6292>FT370000). In other words, the map of basal R1b-V88* below might include any of the undifferentiated clades currently described upstream of R-V88. Furthermore, the reported data did not include much of the Arabian Peninsula, and the data as a whole is an amalgam of new and previous studies with varied coverage of SNPs.
Frequency maps of the main trans-Saharan sub-clades of R1b-V88. For each map, the relative frequencies (percentages) are reported to the right. Image modified from D’Atanasio et al. (2018).*Maps represent the range of the following subclades in FTDNA: V88>FGC21056>FGC21027>FGC21028>FGC21009>V1589. V88>FGC21056>FGC21027>FGC21028>FGC21009>V1589>FT57612>BY39607 (equivalent to V4759). V88>FGC21056>FGC21027>FGC21028>FGC21009>V1589>FT57612>FT219266>BY14102>V69.
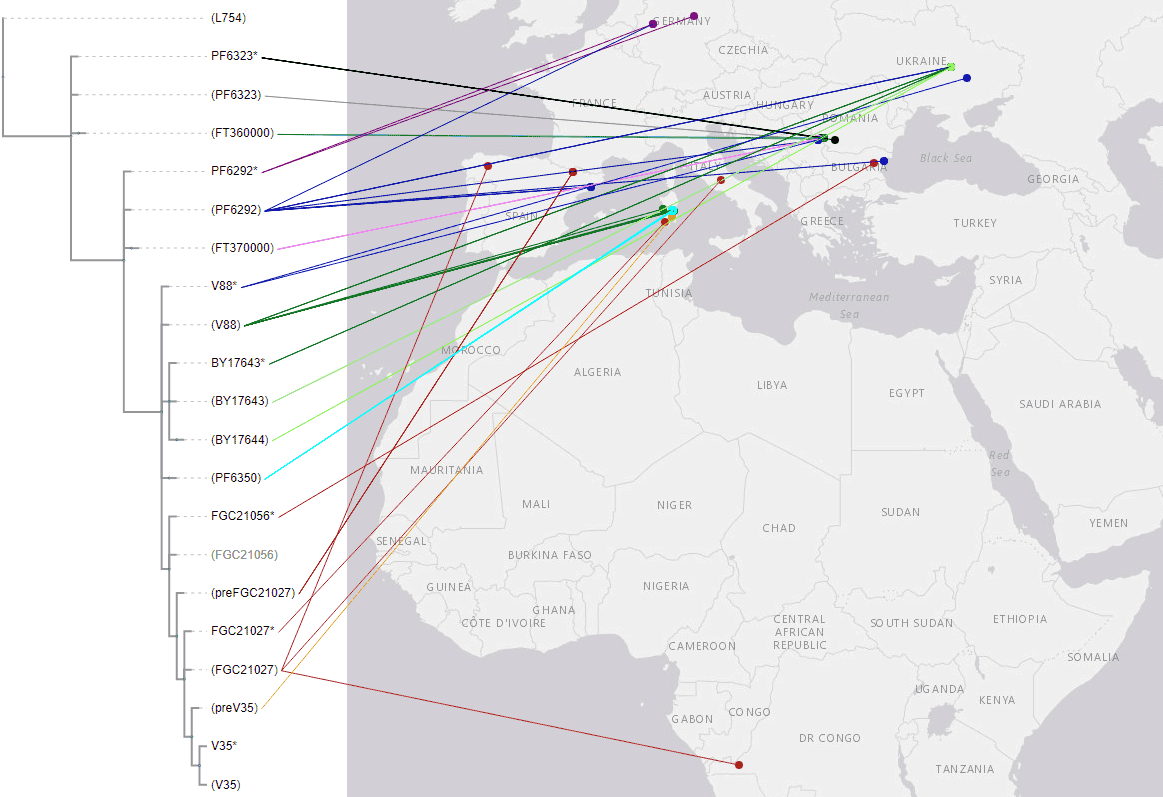
2)My own TreeToM phylogeographic maps of clades based on YFull’s reported Y-names was revealing, but also wrong in some key aspects, and incomplete in others. I have checked some samples I believed could be relevant, and you can see below how the new map with the phylogenetic tree looks like. It is now even more evident at first sight that the core of this haplogroup’s expansion (including all subclades at least up to FGC21056) lies in the Balkan Neolithic, with Cardial/Epicardial samples and later derived ones of the Central or Western Mediterranean representing likely sinks of a South-Eastern European source.
NOTE. For more information on each individual sample, refer to the ancient DNA dataset; for chronologically layered samples, see the ArcGIS WebMap. I haven’t checked out some of the samples for FTDNA equivalents, though, so they might be below their current clade, or even wrongly assigned.
TreeToM phylogeographic map of hg. R1b-PF6323 among ancient samples. See interactive version.
As far as the mtDNA is concerned, both Oman BA samples taken together point to a common H2a2a1. This haplogroup is found much earlier in an Iberomaurusian individual from Taforalt (ca. 20000 BC), and it is also found later overall in Europe, after the Neolithic expansion. Upper clades H2a2 and H2a2a (potentially including this one) are also found in Anatolia Neolithic and as far east as Turan.
Also relevant for this discussion is the reference in Gad et al. (2020) to the Royal Egyptian male lineage of haplogroup R1b that was passed from the grandparent (Amenhotep III) to the father (KV55, Akhenaten) to the grandchild (Tutankhamun) during the 18th Dynasty (in the 14th to 13th century BC).
NOTE. An old STR report for this line suggested it might have belonged to the M269 tree (in fact, a P312>U152 subclade), which seems compatible with the purported origin of the dynasty after the Hyksos period, and with the presence of R1b-L23 in the Middle East during the Bronze Age. However, SNP inference based on STRs is well known to be unreliable, and particularly biased toward the available commercial sampling, which is in turn heavily shifted toward haplogroups present among European testers.
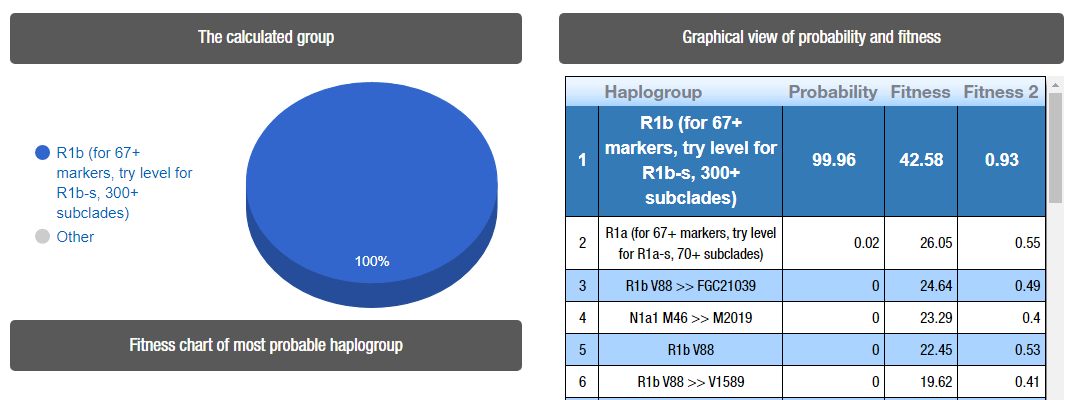
#EDIT (2/14/2021): The newly reported STRs show a more likely R1b-V88, based on the results below taken from Nevgen (Gad et al. 2020 use Athey):
Caption from Gad et al. (2020): Maternal and Paternal Lineages in King Tutankhamun’s Family.
For those who would like to test the STR prediction for Akhenaten, introduce the following data in the box: 13,24,14,11,11-14,0,0,10,13,13,30,16,0-0,0,0,0,14,19,0,0-0-0-0,0,11,0-0,15,0,0,0,0-0,0,12,0,0,0-0,0,0,0,0,0,0,0,0-0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,23
Caption of Nevgen’s prediction for the haplogroup of Akhenaten.
Another ancient V88 sample has been reported from Africa in Wang et al. Sci. Adv. (2020), KIN004 from Kindoki in the modern DRC (ca. AD 1720), of subclade R1b-V88>FGC21056>FGC21027, hence possibly forming part of the Bantu expansion, or alternatively as a recently acculturated Hausa or Fulani lineage.
Neolithic expansion
It is well known by now that the expansion of the so-called “Neolithic package” with Early European Farmers of NW Anatolia Neolithic-related ancestry was not – as it could have been initially thought – a mere spread of a single community from Anatolia throughout Europe. The current data shows that the expansion of EEF ancestry was probably the result of the stepped replacement of demographically stronger agricultural communities by hunter-gatherer groups, marked by different Y-DNA bottlenecks in mainly east-to-west and south-to-north waves of expansion. This is reflected in the discontinuous spread of distinct communities marked by different cultural traits and showing a variable degree of continuity in terms of genome-wide ancestry, often accompanied by completely different Y-chromosome haplogroup bottlenecks.
It is unclear how the majority of African R1b-PF6323 clades relate to the newly developed tree by FTDNA, since the number of testers is not comparable to those from Europe. So far, different clades related to the prevalent African ones have been found in Neolithic and later samples from Central Italy, Sardinia, and North-Eastern Iberia, which made me always think about a Central Mediterranean route into Northern Africa. However, North-Western Africa has been reported more than once already to show continuity in terms of genome-wide ancestry and lineages, and basal clades are prevalent among Mesolithic and Neolithic Balkan populations, which strongly suggests that South-Eastern Europe was the hotspot of R1b-PF6323 by the time the Neolithic package expanded from Anatolia into Europe, and continued to be so during the Chalcolithic. Therefore, the Eastern Mediterranean looks like a more plausible source of the migration of clades currently found in Africa and the Middle East.
With the current poor archaeological description, it is impossible to know whether there might have been fitting Levant / North-Eastern African Neolithic migration waves reaching as far to the south-east as the Omani coasts. These Oman BA samples are probably later than the nomadic and semi-nomadic Omani Neolithic populations that moved through the inner regions and along the coast, and belong thus to the end of the Late Neolithic, characterized by dry climate and the development of sedentary life by the then ‘indigenous’ population on the coasts, featuring Ra’s al-Hamra Facies around Muscat and Sur. The samples might even belong to the later transition phase into Bandar-Jissah Facies, where people used metal as well as stones in the manufacture of their tools (Al-Jahouri, 2008), because it is not clear why the authors labelled them Oman_BA instead of Oman_LN.
My wild guess based on those Ra’salhamzian samples would be that, if they belong to the earlier Neolithic expansions of hg. R1b-PF6323 (and not to Palaeolithic ones of R1b-L754*), they might reflect an early migration wave of an Anatolia Neolithic-like South-Eastern European population to the Levant and/or North-Eastern Africa, which winded up in the Omani coasts after a period of migrations of (semi-)nomadic populations through Arabia. Its current distribution could then be similar to the expansion of hg. T, whose history in the Levant, with back and forth Neolithic migrations throughout the Mediterranean and the Middle East, is not at all clear.
Of course, it could also be that the Oman BA subclades are more related to Villabruna than to SE European PF6323. Or, maybe they are not even R1b (and neither K, P, F, CT…).
It is good practice to be registered
and logged in to comment.
Please keep the discussion of this post on topic. Civilized discussion. Academic tone.
For other topics, use the forums instead.
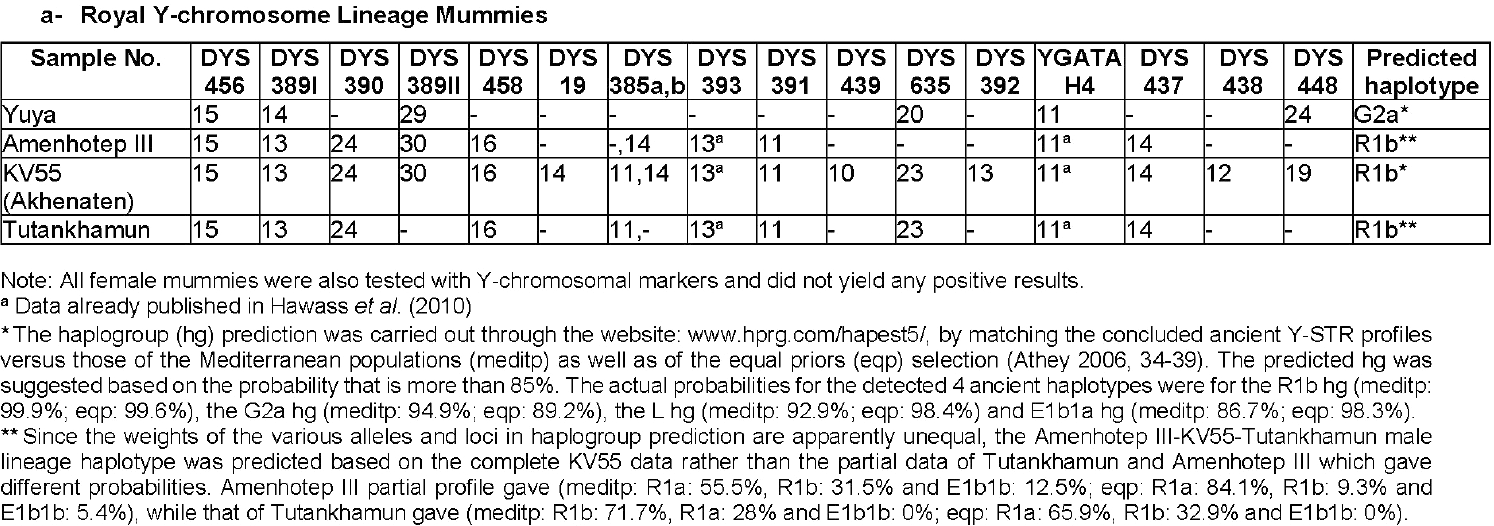



{kind=link}
{kind=link}