Artículo de libre acceso Rastreando cinco milenios de domesticación del caballo con una serie temporal extensa de genomas ancestrales, por Fages et al. Cell (2019).
Extractos interesantes (traducción automática, énfasis mio):
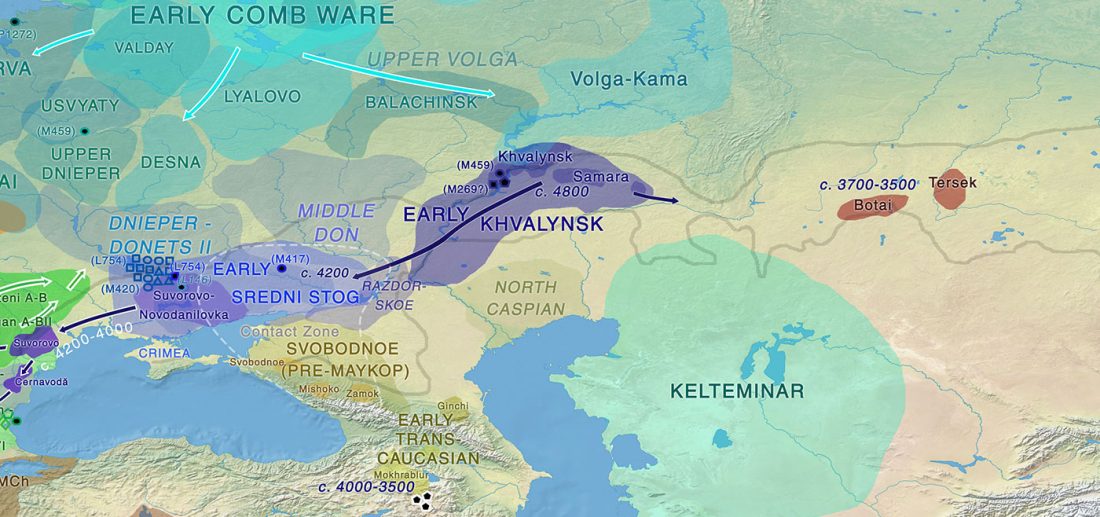
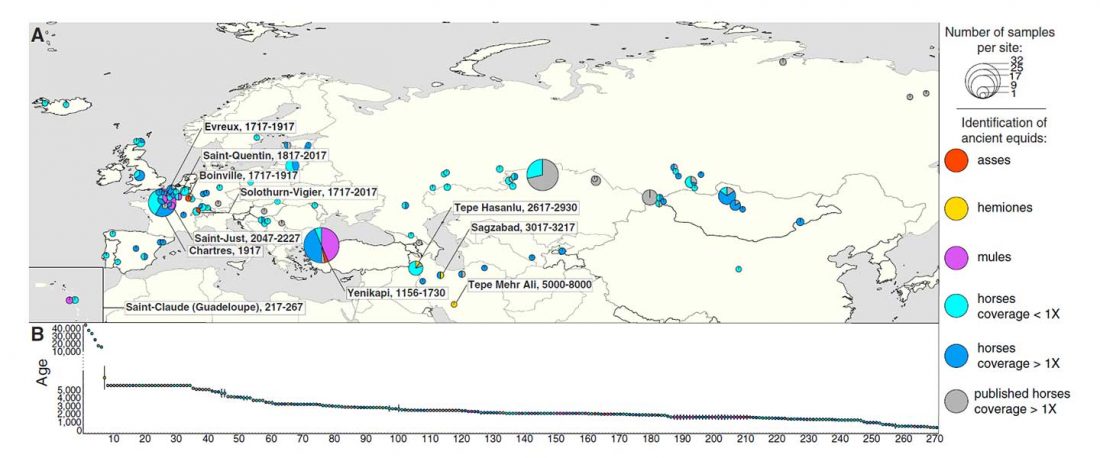
… Read the rest “Yamna la fuente más probable del caballo doméstico; el linaje más próximo, del campaniforme oriental”La evidencia arqueológica más temprana de ordeño, aprovechamiento y acorralamiento de caballos se encuentra en la cultura Botai de ,5005,500 años de las estepas de Asia Central (Gaunitz et al., 2018, Outram et al., 2009; véase Kosintsev y Kuznetsov, 2013 para discusión). Sin embargo, los caballos tipo Botai no son los ancestros directos de los domesticados modernos sino los caballos de Przewalski (Gaunitz et al., 2018). Por lo