Artículo de libre acceso Rastreando cinco milenios de domesticación del caballo con una serie temporal extensa de genomas ancestrales, por Fages et al. Cell (2019).
Extractos interesantes (traducción automática, énfasis mio):
La evidencia arqueológica más temprana de ordeño, aprovechamiento y acorralamiento de caballos se encuentra en la cultura Botai de ,5005,500 años de las estepas de Asia Central (Gaunitz et al., 2018, Outram et al., 2009; véase Kosintsev y Kuznetsov, 2013 para discusión). Sin embargo, los caballos tipo Botai no son los ancestros directos de los domesticados modernos sino los caballos de Przewalski (Gaunitz et al., 2018). Por lo tanto, el origen genético de las domesticadas modernas sigue siendo polémico, con sugerencias de candidatos en las estepas Pontic-Caspian (Anthony, 2007), Anatolia (Arbuckle, 2012, Benecke, 2006) e Iberia (Uerpmann, 1990, Warmuth et al., 2011). Independientemente de los orígenes de la domesticación, se sabe que el genoma del caballo ha sido remodelado significativamente en los últimos ,32,300 años (Librado et al., 2017, Wallner et al., 2017, Wutke et al., 2018). Sin embargo, en qué momento y en qué contexto (s) se produjeron tales cambios sigue siendo en gran parte desconocido.
Para aclarar los orígenes de los caballos domésticos y revelar su posterior transformación por civilizaciones ecuestres pasadas, generamos datos de ADN de 278 subfósiles equinos con edades que abarcan los últimos seis milenios (n = 265, 95%) (Figuras 1A y 1B; Tabla S1; Métodos STAR). El contenido de ADN endógeno fue compatible con la secuenciación económica de 87 genomas de caballos nuevos con una profundidad de cobertura promedio de 1.0 a 9.3 veces (mediana = 3.3 veces; Tabla S2). Esto más que duplica el número de genomas de caballos antiguos hasta ahora caracterizados. Con un total de 129 genomas antiguos, 30 genomas modernos y nuevos datos a escala genómica de 132 individuos antiguos (0,01 a 0,9 veces, mediana = 0,08 veces), nuestro conjunto de datos representa la serie temporal más grande a escala genómica publicada para un Organismo no humano (Tablas S2, S3 y S4; Métodos STAR).
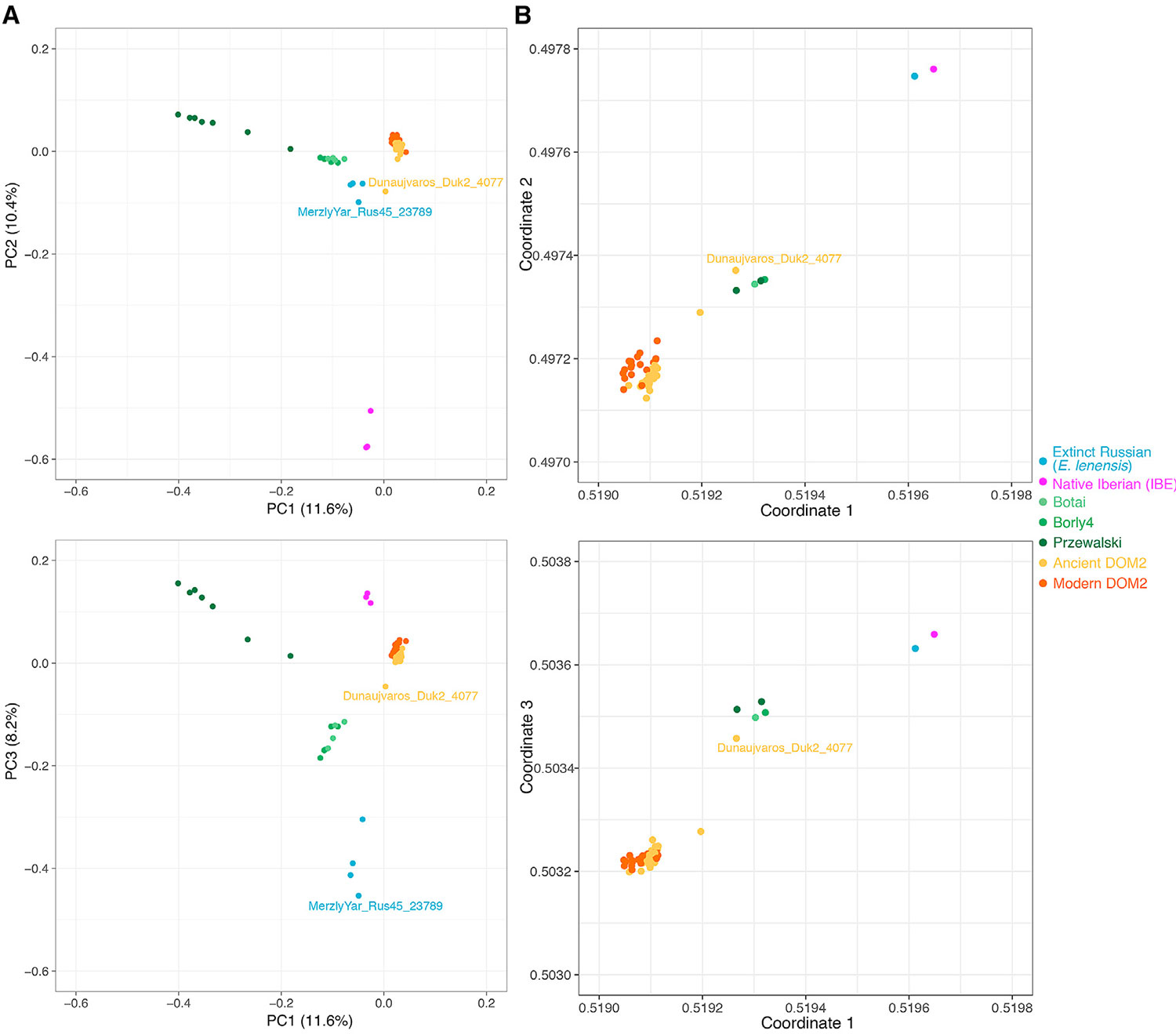
(A) Análisis de componentes principales (PCA) de 159 genomas de caballos antiguos y modernos que muestran una profundidad de cobertura promedio de al menos 1 vez. La estructura genética general se muestra para los tres primeros componentes principales, que resumen el 11.6%, 10.4% y 8.2% de la variación genética total, respectivamente. Los dos especímenes MerzlyYar_Rus45_23789 y Dunaujvaros_Duk2_4077 discutidos en el texto principal están resaltados. Consulte también la Figura S7 y la Tabla S5 para obtener más información.
(B) Visualización de las afinidades genéticas entre individuos, como lo revela el algoritmo struct-f4 y las permutaciones de 878,475 f4. El cálculo de f4 se condicionó a las transversiones de nucleótidos presentes en todos los grupos, y las muestras se agruparon como en los análisis TreeMix (Figura 3). En contraste con la PCA, las permutaciones f4 miden la deriva genética a lo largo de las ramas internas. Por lo tanto, es más probable que revelen una subestructura de población antigua.
Descubriendo dos linajes de caballos divergentes y extintos
Los caballos domésticos y de Przewalski son los únicos dos linajes de caballos existentes (Der Sarkissian et al., 2015). Otro linaje fue identificado genéticamente a partir de tres huesos que datan de ∼43,000–5,000 años atrás (Librado et al., 2015, Schubert et al., 2014a). Mostró afinidades morfológicas a una especie de caballo extinto descrita como Equus lenensis (Boeskorov et al., 2018). Ahora encontramos que este linaje extinto también se extendió al sur de Siberia, luego del análisis de componentes principales (PCA), la agrupación filogenética y f3 de un grupo de spec 24,000 años de la República de Tuva dentro de este grupo (Figuras 3, 5A y S7A). Este nuevo espécimen (MerzlyYar_Rus45_23789) contiene un ADNmt extremadamente divergente que solo se encontró en las islas de Nueva Siberia hace unos ,233,200 años (Orlando et al., 2013) (Figura 6A; Métodos STAR) y está ausente de los tres huesos previamente secuenciados. Esto sugiere que un linaje de caballos fantasma divergente contribuyó a la ascendencia genética de MerzlyYar_Rus45_23789. Sin embargo, el momento y la ubicación del contacto genético entre E. lenensis y este linaje fantasma siguen siendo desconocidos.
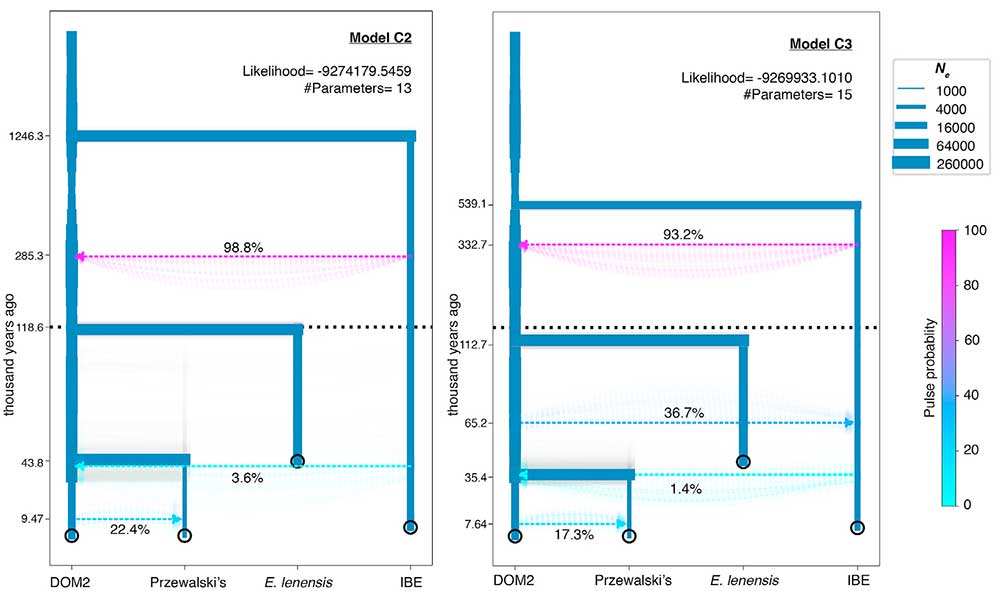
Modelado de demografía y mezcla de linajes de caballo extinto y remanente
Las reconstrucciones filogenéticas sin flujo genético indicaron que la EIB se diferenciaba antes de la divergencia entre DOM2 y los caballos de Przewalski (Figura 3; Métodos STAR). Sin embargo, permitir un borde de migración en TreeMix sugirió afinidades más cercanas con un solo espécimen DOM2 húngaro del 3er milenio BCE (Dunaujvaros_Duk2_4077), con una contribución genética extensa (38.6%) de la rama ancestral a todos los caballos (Figura S7B). Esto, y el cromosoma IBE Y extremadamente divergente (Figura 6B), sugieren que una población fantasma divergente pero no identificada podría haber contribuido a la composición genética de la EIB.
Rechazando la contribución ibérica a los modernos domesticos
Las secuencias genómicas de cuatro especímenes de IBE de 4,800 a 3,900 años de antigüedad caracterizados aquí nos permitieron aclarar los debates en curso sobre la posible contribución de Iberia a la domesticación de caballos (Benecke, 2006, Uerpmann, 1990, Warmuth et al., 2011). El cálculo de la llamada relación fG (Martin et al., 2015) proporcionó un límite mínimo para la contribución de la OIE a los miembros de DOM2 (Cahill et al., 2013) (Figura 7A). El máximo de dicha estimación se encontró en el espécimen Húngaro Dunaujvaros_Duk2_4077 (∼11.7% –12.2%), consistente con su agrupación TreeMix con IBE cuando se permite un borde de migración (Figura S7B). Este espécimen se sugirió previamente para compartir la ascendencia con una población aún no identificada (Gaunitz et al., 2018). El cálculo de la estadística f4 indica que esta población no está relacionada con E. lenensis sino con la EIB (Figura 7B; Métodos STAR). Por lo tanto, la OIE o los caballos estrechamente relacionados con la EIB, contribuyeron con la ascendencia de los animales que se encuentran en un centro comercial de la Edad del Bronce Temprano en Hungría desde el 3er milenio BCE. Esto podría indicar que hubo un intercambio de caballos a larga distancia durante el fenómeno del vaso campaniforme (Olalde et al., 2018). El límite mínimo de la F G para la contribución de la EIB a un caballo español de la Edad del Hierro (ElsVilars_UE4618_2672) seguía siendo importante (~ 9.6% –10.1%), lo que sugiere que una influencia genética de la EIE persistió en Iberia hasta al menos el siglo VII aC en un contexto doméstico. Sin embargo, las estimaciones de fG fueron más limitadas para casi todos los caballos antiguos y modernos investigados (mediana = ~ 4.9% –5.4%; Figura 7A).
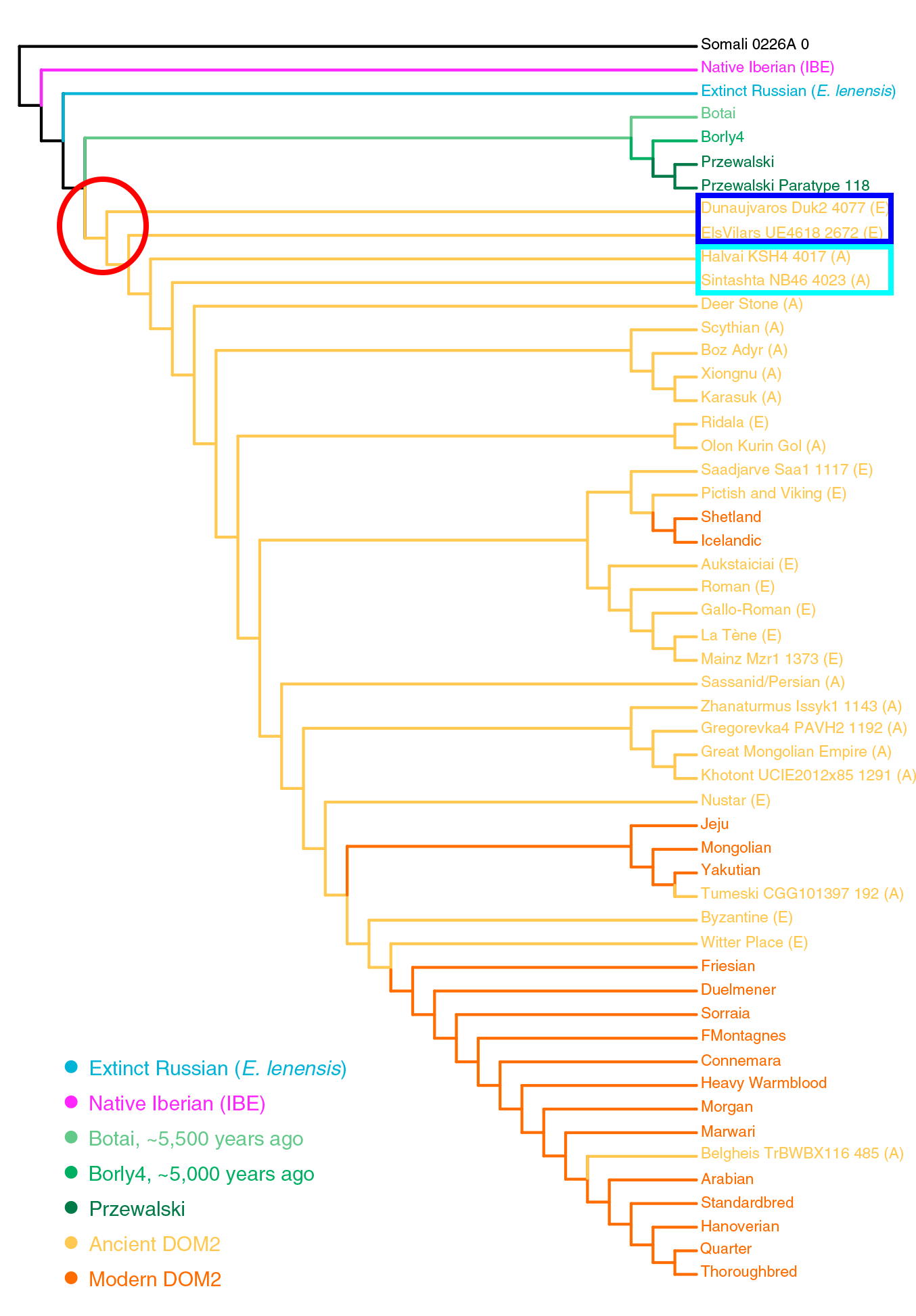
Caballos de la Edad del Hierro
La diversidad de nucleótidos del cromosoma Y (π) disminuyó de manera constante en ambos continentes durante los últimos ∼2,000 años, pero descendió a los niveles actuales solo después de 850–1,350 CE (Figuras 2B y S2E; métodos STAR). Esto es consistente con el predominio de un haplogrupo oriental de ½ a 700 años en la mayoría de los sementales modernos (Felkel et al., 2018, Wallner et al., 2017). Nuestros datos también indican que la creciente influencia de las líneas de sementales específicas post-Renacimiento (Wallner et al., 2017) fue responsable de una caída de 3.8 a 10.0 veces en la diversidad del cromosoma Y.
Luego, calculamos las estimaciones del π del cromosoma Y en culturas pasadas representadas por un mínimo de tres machos para aclarar los contextos históricos que más impactaron la diversidad del cromosoma Y. Esto confirmó la trayectoria temporal observada anteriormente, ya que los caballos bizantinos (287–861 dC) y los caballos del Gran Imperio Mongol (1,206–1,368 dC) mostraron una diversidad limitada pero mayor que la moderna. Caballos de la Edad de Bronce de la cultura de Piedras de Ciervo en Mongolia, caballos medievales de Aukštaičiai de Lituania (siglos noveno a décimo de la Era Común), y Caballos de la cultura Pazyryk de los escitas durante la Edad del Hierro mostraron niveles de diversidad similares (0.000256-0000267) (Figura 2A). Sin embargo, la diversidad fue mayor en los caballos de La Tène, romanos y galorromanos, donde las proporciones de π de Y a autosómica fueron cercanas a 0.25. Esto contrasta con los caballos modernos, donde la marcada selección de líneas específicas conduce a relaciones π a autosómicas sustancialmente por debajo de 0.25 (0.0193-0.0396) (Figura 2A). Las proporciones de cerca de 0.25 Y a autosomal π encontradas en los caballos La Tène, romanos y galorromanos sugieren estrategias de reproducción que incluyen un éxito reproductivo uniforme entre sementales o un éxito reproductivo igualmente sesgado en ambos sexos (Wilson Sayres et al. 2014).
El término linaje se usa en este documento, como en muchos otros en genética, definido por una ascendencia específica. Mantengo esa nomenclatura a continuación. No debe confundirse con los “linajes” o “líneas” que se refieren a los haplogrupos del cromosoma Y (o ADNmt).
Apoyando la naturaleza “arcaica” de los caballos húngaros del campaniforme que se expanden desde las estepas póntico-cáspias son:
- Entre las líneas del cromosoma Y, el grupo común formado por Botai-Borly4 (estrechamente relacionado con DOM2) , caballos escitas de Aldy Bel (Arzhani), caballos de la Edad del Hierro de Estonia (Ridala), caballos de la cultura Xiongnu (Uushgiin Uvur), y caballos romanos de Autricum (actual Chartres).
- Entre las líneas de ADNmt, el grupo común formado por las muestras de Botai ,LebyazhinkaIV NB35, y diferentes domesticados euroasiáticos, incluidos muchos de los antiguos europeos occidentales, revelando una probable expansión de ciertos subclados al este y oeste con la cultura Repin.
(…)(…) DOM2 contribuyó con un 22% al ancestro de los caballos de Przewalski ca. 9.47 kya, sugiriendo el óptimo holoceno, en lugar de la cultura botai eneolítica (∼5.5 kya), como un período de contacto con la población. Esta introgresión previa a Botai podría explicar la topología del cromosoma Y, donde se informó que los caballos de Botai portaban dos haplogrupos segregantes diferentes: uno ocupaba una posición basal en la filogenia, mientras que el otro estaba estrechamente relacionado con DOM2. Sin embargo, se sabe que se produjeron múltiples pulsos de mezcla a lo largo de la divergencia de DOM2 y el linaje Botai-Borly4, que incluye un 2,3% de la contribución post-Borly4 al DOM2 y una intogresión DOM2 más reciente del 6,8% en los caballos de Przewalski (Gaunitz et al., 2018). Los parámetros del Modelo C2 acomodan todo esto como un solo pulso de mezcla, probablemente promediando las contribuciones de todos estos eventos múltiples
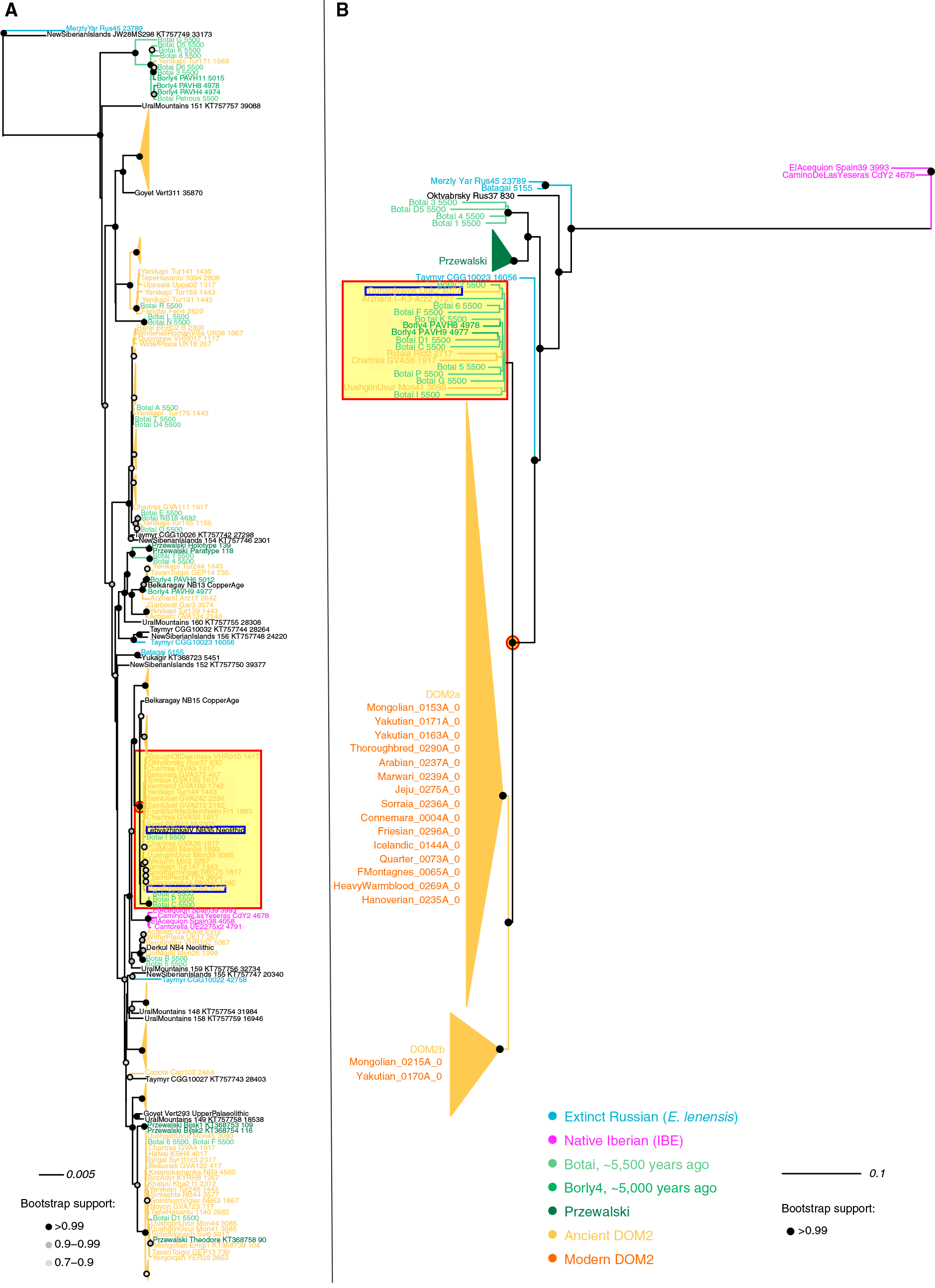
(A) Árbol de máxima verosimilitud que recorre las relaciones filogenéticas entre 270 genomas mitocondriales.
B) El mejor árbol de probabilidad máxima del cromosoma Y (modelo de sustitución GTRGAMMA) excluyendo el grupo externo. Los soportes de nodo se indican como fracciones de 100 pseudorreplicados de arranque. No se muestran los soportes Bootstrap inferiores al 90%. La raíz se colocó en el punto medio del árbol. Consulte también la Tabla S5 para obtener información sobre el conjunto de datos.
Imagen modificada del papel, que incluye un cuadrado rojo en grupos arcaicos que contienen la muestra húngara y un círculo rojo alrededor del semental ancestral más común y la yegua de las estepas Pontic-Caspian.
El documento no puede ofrecer una imagen detallada de la domesticación de caballos en la Prehistoria, pero es otro paso más para mostrar cómo Repin / Yamna es la fuente más probable de expansión de los caballos domésticos en Eurasia. Aún más interesante, los colonos Yamna en Hungría probablemente expandieron un antiguo linaje de ese caballo al mismo tiempo que se expandieron con la cultura campaniforme clásica. Se encuentran así paralelos notables entre:
- el linaje de la muestra Dunaujvarus Duk2 4077 y la expansión del indoeuropeo noroccidental (ca. 3000-2500 aC).
- el linaje de las muestras Halvai KSH4 4017 y Sintashta NB46 4023 y la posterior expansión protoindoirania (ca. 2500-2000 aC).
La expansión de una antigua línea de caballos domésticos relacionados con yamna Hungaría / campaniforme oriental parece ser confirmada por la muestra preibérica de Vilars I, Els Vilars4618 2672 (ca. 700-550 aC), probablemente de ascendencia de campaniformes ibéricos, mostrando un linaje más antiguo que los indoiranios, que más tarde reemplazaron a la mayoría de las líneas paternas europeas.
NOTA.Para contactos conocidos entre Yamna y protocampaniformes justo antes de la expansión del campaniforme oriental, vea una publicación reciente sobre los grupos yamna de vanguardia hasta Sevilla .
Los hallazgos del artículo confirman la expansión del caballo primero (y principalmente) a través del bioma de la estepa, imitando primero siguiendo la expansión de los protoindoeuropeos y luego siendo reemplazados gradualmente (o no tan gradualmente) por líneas traídas a Europa por culturas de la estepa especializadas en equitación, durante sus expansiones hacia el oeste en la Edad de Bronce, la Edad de Hierro y el medievo temprano. La expansión también se correlaciona bien con la propagación conocida de la tracción animal y el pastoreo antes del año 2000 aC:
EDITADO (3 Mayo 2019): un recordatorio reciente de estos desarrollos paralelos por parte de David Reich en Insights in visiones de las expansiones de idiomas con el ADN antiguo:
- La expansión de la cultura yamna hacia el oeste “con caballos y carros”, encontrándose una ascendencia más homogénea en los europeos modernos debido a migraciones posteriores desde el este (y el norte):
- “Descendientes” de yamna (una vez que la cultura ya estaba “muerta”), expandiéndose hacia el este principalmente con la ascendencia de cerámica cordada :
- The genetic and cultural barrier of the Pontic-Caspian steppe – forest-steppe ecotone
- Mitogenomes suggest rapid expansion of domesticated horse before 3500 BC
- Origin of horse domestication likely on the North Caspian steppes
- Ancient DNA upends the horse family tree
- Domesticated horse population structure, selection, and mtDNA geographic patterns
- About Scepters, Horses, and War: on Khvalynsk migrants in the Caucasus and the Danube
- Steppe and Caucasus Eneolithic: the new keystones of the EHG-CHG-ANE ancestry in steppe groups
- Domestication spread probably via the North Pontic steppe to Khvalynsk… but not horse riding
- Differing modes of animal exploitation in North Pontic Eneolithic and Bronze Age Societies
DR: inference is that two major migrations: farmers from Anatolia, followed by steppe pastoralists. Who are they? They took horses and wagons and spread. See rapid 90% pop turn over in Britain. Similar timing in Iberia, but a bit less turnover, and more period of overlap
— Joshua G. Schraiber? (@jgschraiber) May 2, 2019
DR: spread of steppe ancestry to the east likely a result of spread of yamnaya descendents, since yamnaya were dead. High genetic similarity to corded ware people.
— Joshua G. Schraiber? (@jgschraiber) May 2, 2019
Otro artículo reciente de acceso abierto sobre la domesticación de caballos es The horse Y chromosome as an informative marker for tracing sire lines, por Felkel et al. Scientific Reports (2019).