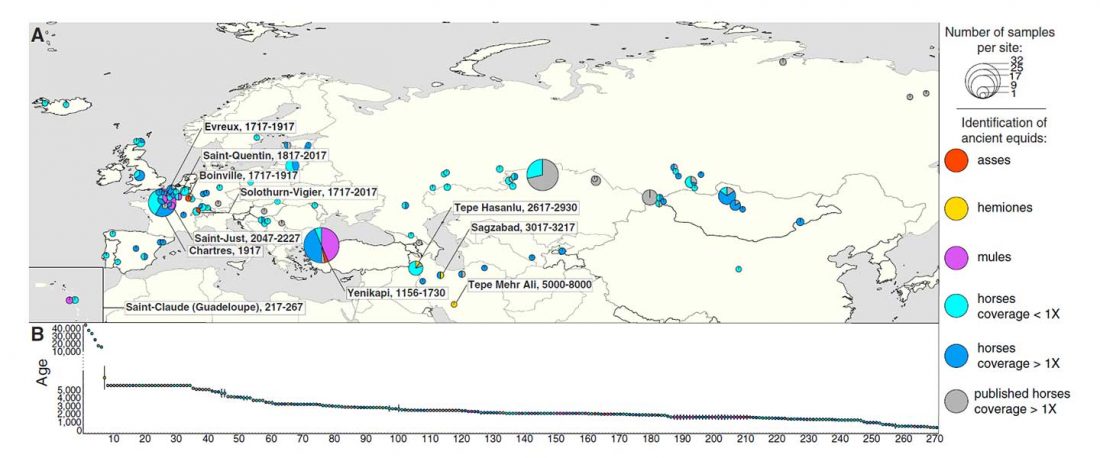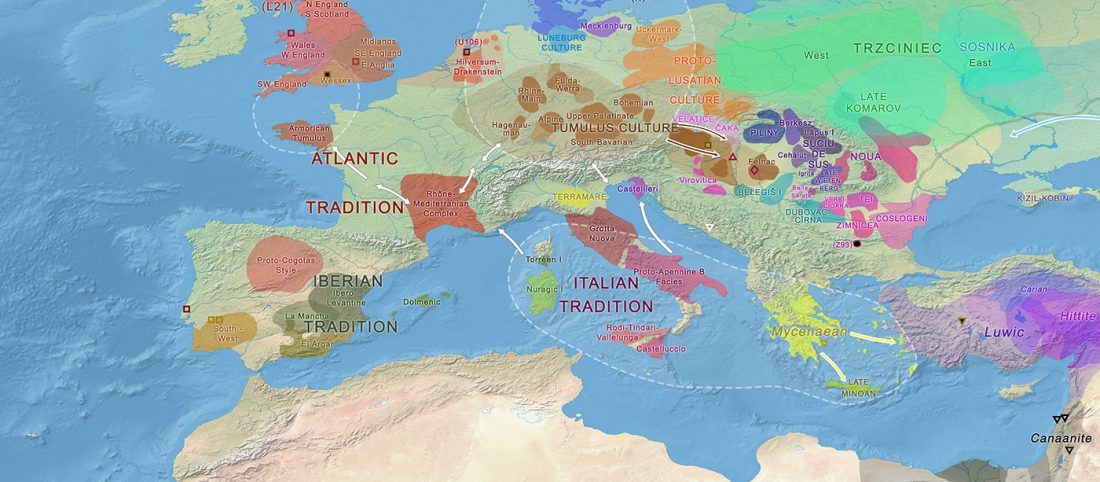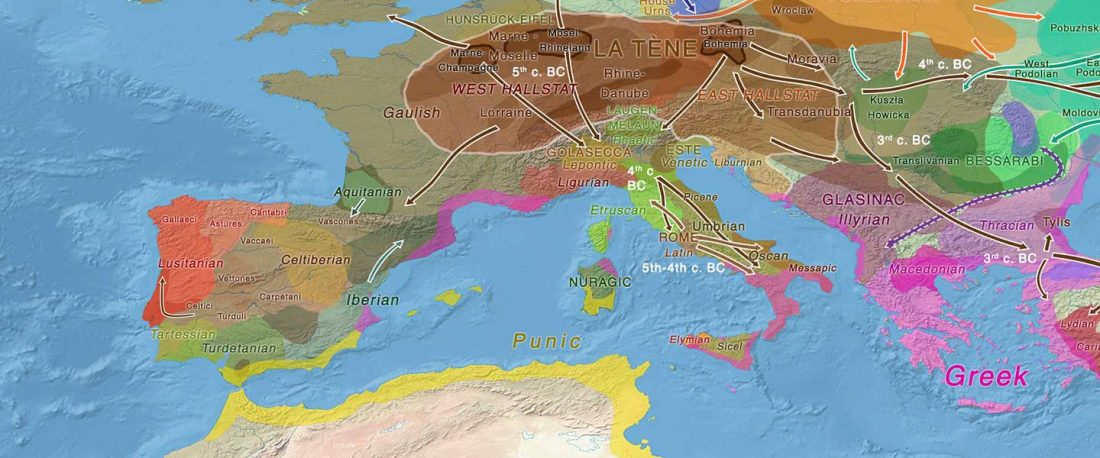
Los datos de ADN antiguo de Iberia publicados por Olalde et al. (2019) son interesantes por varios motivos, pero aún tengo la impresión de que los autores – y por consiguiente muchos lectores – se centraron en información poco relevante acerca de movimientos poblacionales muy recientes, o incluso destacaron los detalles menos importantes relacionados con eventos históricos.
Ya he escrito sobre la relevancia de estos hallazgos para la cuestión indoeuropea en una opinión inicial (inglés), luego en un post más detallado sobre sus consecuencias (inglés), luego sobre la llegada de lenguas célticas con el hg. R1b-M167, y … Read the rest “Indoeuropeos preceltas descendientes de campaniformes ibéricos de haplogrupo R1b-P312”